Welcome to the next issue of the Seaweed for Sceptics series. The first post of the series explored the rough origin of the red algae, showing the genetic impariment that these seaweed suffer as a result of their extreme origins. In this second post we shall focus on one of these weird connections that crop up between branches of the tree of life: seaweed and ammonoids.
Now presenting: the ammonoids
Ammonoids are popular fossil cephalopods, originating in the lowermost Devonian and dying out at the base of the Paleocene (Danian). The typical ammonoid sported a monomorph shell shape, i.e., had a spiral shell, elegantly coiling in a single plane, with all whorls in tangential or overlapping contact.
Interesting enough, a number of ammonites depart from the arquetypical image of a single coil; with puzzling coiling patterns that involve elbows, uncoiling, twisting in many planes… The ammonites that depart from the monomorph shape are called heteromorphs. You can find pictures exploring the picture galleries from here, here or here.
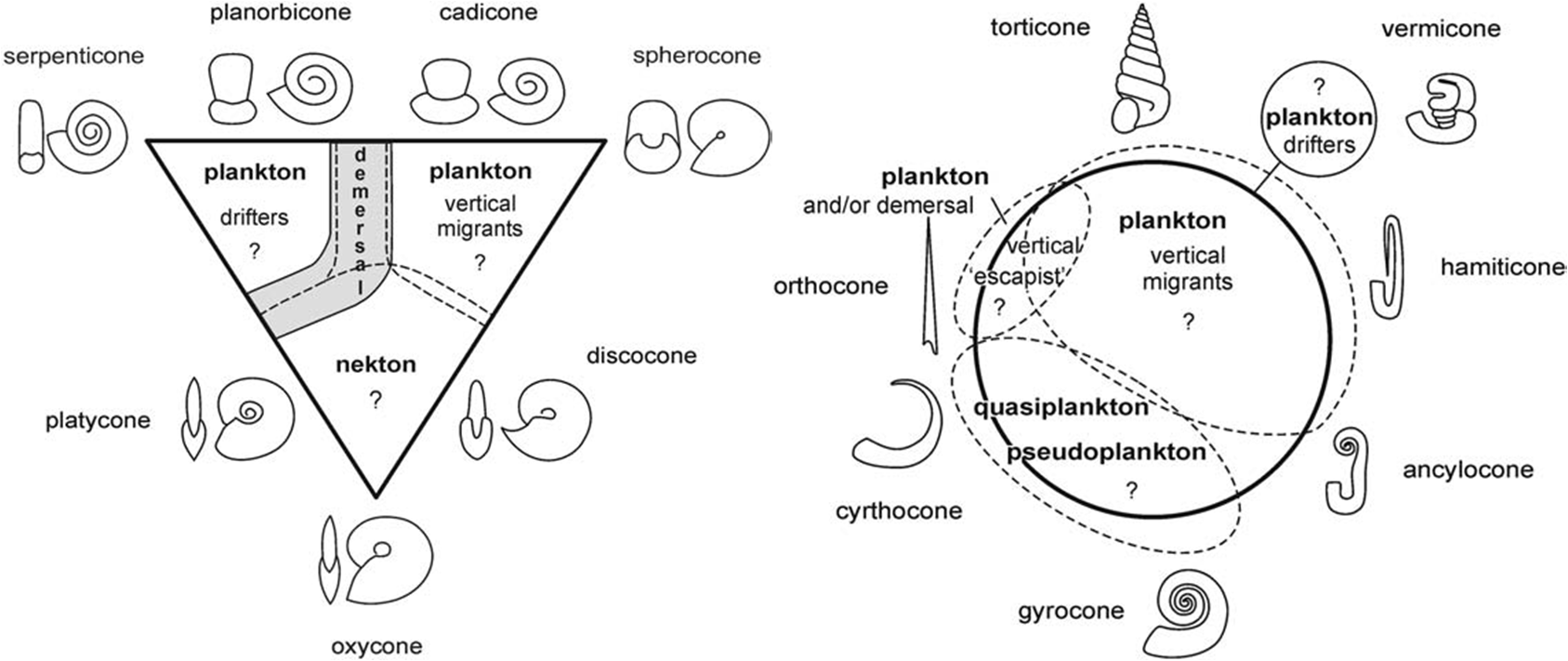
Monomorphy and heteromorphy refer exclusively to shell shape, they have no phylogenetic meaning. Heteromorphs don’t form a natural group (neither do monomoprhs). Heteromorph forms have evolved repeatedly and independently from monomorphs in separate lineages.
In this super-brief introduction about ammonoids I’ve focused on the shell for a reason: it’s the only thing we have. They left almost no fossil aside from their shell, the soft-tissue anatomy remains largely unkown. Thus, the taxonomy, classification and ecological interpretations rely (almost) solely upon the structure and form of the shells.
Ammonoid ecology: many speculations (and seaweed?)
Inferring the ecology, behaviour or habitat of an ammonoid using the shell alone is no easy task. There has been a considerable amount of debate, and ecological interpretations remain quite speculative. You know what they say: In the absence of proper data, speculate wildly.
Given the wide diversity of coiling patters, a common life style seems unlikely. Most monomorphic ammonoids are considered to have been pelagic (roughly equally divided between drifters, swimmers and vertical migrants), with a few bottom-feeders.
There is room for seaweed in this ecological conundrum. Floating and benthic algal mats have been proposed as an habitat for both monomorphs and heteromorphs, as well as possible spawning places. Association with algal mats may have reduced vulnerability to predation.
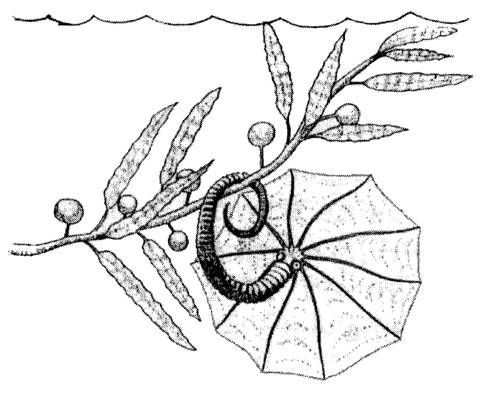
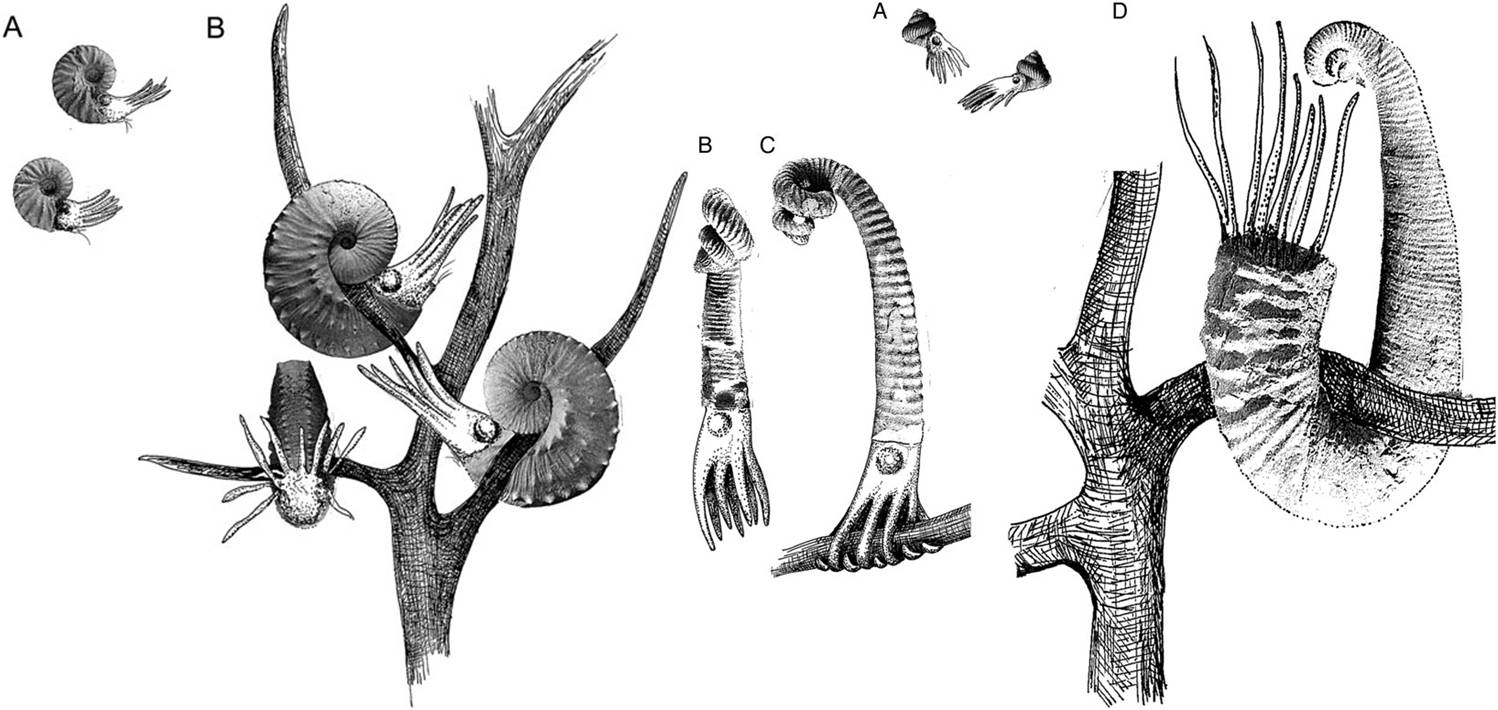
In 2014, a certainly creative link was proposed between T-C heteromorphs* (belonging to the suborder Ancyloceratina) and seaweed (namely the Phaeophyta): adults were stationary, hooked or clipped to algal stipes and branches.
* Terminal-countdown heteromorphy (T-C for short) is most often represented in ammonoids by an U-shaped body chamber. T-C heteromorphs combine determinate growth with the development of novel form.

Branches are an ammonoid’s best friend
Various features of the adult shell are consistent with the stationary adult hypothesis: a weaker (sometimes worn) ribbing pattern on the inner surface of the hook, asymmetry of adult habitation chamber, lack of epizoans (parasitic animals glued to the external surface of the shell) and variability in adult shell shape.
This T-C morphology has been shown to be evolutionary convergent, yet temporally constrained essentially to the Cretaceous. The convergent evolution of this fancy coiling by a number of lineages simultaneously and independently of one another lends support to the idea that there was an ecological shift going on at that time.
This shift could be an adaptative response to radiations of algal macrophytes (forming kelp-like forests), following the adoption of a new reproductive strategy, with a sedentary adult female phase in which the eggs were brooded while the shell was hooked.
An stationary brooding phase has some advantages. Females would have been able to channel more resources into egg production (since less energy is spent in active swimming), the seaweed provided concealment from pelagic predators, and as seaweed inhabit warm and productive areas, egg development would have been faster, and hatchlings would have had more resources (compared to offshore habitats).
The right type of seaweed
What kind of branches were these fancy heteromorphs attached to? The best candidates seem to be our good friends, the seaweed. Branching colonial animals, such as bryozoans, crinoids and corals were abundant in the Late Cretaceous oceans… but the calcified branches would have caused greater wearing of the ribbing pattern on the inner side of the hook. Following a similar reasoning, calcified seaweed can be ruled out (Rhodophyta and some green algae).
We are left with the brown algae* (Phaeophyta). The Phaeophyta are a distinct group of eukaryotic algae with approximately 1.800 modern species. They exhibit quite a variety of life cycles and reproductive strategies. Most of them are found at sea (just 4 genera with freshwater species), with a worldwide distribution, yet they are predominant only in cold waters. Some of these algae are well known as kelp (most of them belong to the order Laminariales), big, fleshy macroalgae with robust stems and stipes. Supposedly the stipes were able to hold the curved shell of the ammonoid.
* Calcification of the thallus is known for some species of Padina, nevertheless, the fan-shaped structure of this elegant algae makes it unsuitable as a candidate.

The fossil record of the brown algae is quite poor (alas, no calcification means small chances of fossilization). The exact timing of the origin, radiation and diversification of the main brown algae lineages is not known. Various lines of evidence (genetic, geological, palaeo-oceanographic) suggest that brown macroalgae radiated during the Early Cretaceous, becoming more diversified during the Late Cretaceous. These two events of proposed diversification of brown algae coincide with two peals in diversity of T-C heteromorphs, during the Early and Late Cretaceous.
Personally, I find this hypothesis to be creative, bold, and highly speculative (which is something I tend to like), perhaps even bordering on the ludicrous side of speculation. But nevertheless it is thought-provoking and worthy our time to examine. Even it ends up being wrong, it will still have value. All the effort that is put into debunking wrong hypothesis might bring forward better ideas (even if that was not the primary goal).
A few problems can be pointed out. Some heteromorphs were covered in spikes. What use could the spikes serve in the middle of a kelp forest? They make more sense in a planktonic environment (as a way to avoid predation and/or sedimentation). Kelp tends to dominate the intertidal zone, meaning that they remain emerged for some time (they have a special wall composition to prevent dehydration). If the same is true for Cretaceous kelp, would it really have been a suitable habitat for ammonoids?
I can visualize an ammonite getting hooked onto a branch, but I really can’t see it remaining that way. For some reason, I imagine the shells sliding away, drifting away from the stems. The radiation of heteromorph forms could have been fueled by a greater abundance of food, since heteromorphs would have been able to feed in spite of their non-hydrodynamic or inefficient shell shapes (Juárez, J. & Matamales, R., personal communication, 2016)
UPDATE (27/05/2016): Well, the original 2014 article was indeed thought-provoking (told you so!). There has been a reply (here) againt the hypothesis, and a counter-reply defending it (here). Be sure to check the comment section in this post for further discussion on this matter!
If you would like to share any ideas or suggestions, let us now in the comment section below!
References
Arkhipkin, A.I., 2014. Getting hooked: the role of a U-shaped body chamber in the shell of adult heteromorph ammonites. Journal of Molluscan Studies, 80(4), pp.354–364.
Kaplan, P., 2002. Biomechanics as a Test of Functional Plausibility: Testing the Adaptive Value of Terminal-Countdown Heteromorphy in Cretaceous Ammonoids. Abhandlungen der Geologischen Bundesanstalt, 57, pp.181–187.
Lee, R.E., 2008. Phycology 4th ed.
Lukeneder, A., 2015. Ammonoid Habitats and Life History. In C. Klug et al., eds. Ammonoid paleobiology. Topics in Geobiology. Dordrecht: Springer Netherlands, pp. 697–800.
Westermann, G.E.G., 1996. Ammonoid life and habitat. In N. H. Landmann, K. Tanabe, & R. A. Davis, eds. Ammonoid paleobiology. Topics in Geobiology. Springer, pp. 607–707.
If you enjoyed this article do not forget to follow us on Twitter and Facebook !

Congratulations for the detailed and well explained post! 🙂 I know how difficult and challenging it might be to make a summary of this absurdly vast and diverse group in such a little space. Let me elaborate on my opinion on this topic in a (sorry) quite long comment.
Along with the fact that ammonoids would not really be able to get effectively affixed to the stems without gliding away, as you well pointed out, another big flaw for this theory might be that in some species, both sexual dimorphs feature this kind of growth.
For example, the antidimorphs of the genera Moutoniceras or Amorina (and many others) are virtually identical in shape except for their size difference. So why would males need to get hooked to algae stems? To me, it doesn’t quite add up.
On the other hand, we’ve got other dimorphic couples (Acrioceras-Emericiceras is the best example I can come up with right now), that have T-C microconchs (=Acrioceras) and crioconic macroconchs (=Emericiceras). Then you could say ‘yeah, so the micros are females and macros are males’. However big macroconchs are known to dwell and die in shallow, warm platforms, while small microconchs are found in deeper facies. This has been understood as a migration of females in its late life to spawn in warmer and more protected environments. So males would be the T-C and females not!?
Finally, some species might or might not develop partially T-C morphotypes. For example, the genus Crioceratites is usually characterised by crioconic antidimorphs that usually differ only in size. Some particular individuals of certain species of Crioceratites (such as Cr. curnieri, Cr. duvalii, Cr. villersianum, Cr. “majoricensis”, Cr. binelli…) may grow a “pseudo-hook” off a crioconic shape. The same thing may happen with very big macroconchs of Emericiceras…
As you well said, the purely speculative aspect of this work might seem very interesting at first glance. Nevertheless, once one starts to look for small details, the whole theory might just flounder in farfetchedness. With T-C heteromorphs there’s definitely more than meets the eye, since our eye has hitherto just been able to see the shell (and the carbonised tentacles of a baculitid… :-/).
LikeLike
Wow! That was indeed a long and interesting comment! 🙂 I was unaware of the relevance of sexual dimorphism, and I’m frankly puzzled by ammonoids developping “pseudo-hooks”… Thank you for bringing that up! It is quite a challenge to make sense of these animals. If I may ask, how do you picture the heteromorphs? What kind of habitat do you make out for them? Btw, the original 2014 article has sparked a reply and counter-reply, I’ve updated the links at the end of the post.
LikeLike
Hi again,
First of all, after re-reading the mentioned paper, I should point out that “Terminal-Countdown” is for some reason shortened to T-S instead of T-C, as you put it on the post and I put it on my comment (wrongly).
I mentioned and invented this term for “pseudo-hooks” to apply it on very opened hooks, that form as a more or less conspicuous eccentricity in the original crioconic coiling (see genus Subaspinoceras for examples; in fact, they might just be Crioceratites with freaky growing). I can hardly picture those ammonites getting effectively hooked on algae, however being the most probable ancestors of the first truly “hooked” Ancyloceratina (i.e. Paraspinoceras).
How do I picture the heteromorphs? In their juvenile stages, the living habits might have been tremendously varied: benthic in turriconic forms (i.e. Heteroceras), nektic in monomorphs (i.e. Macroscaphites), planktic in crioconic and protancyloconic shapes (i.e. Acrioceras)… Those living styles are vastly explored in literature and I don’t see why they wouldn’t be as supposed. The part where it uncoils and forms the descending shaft would correspond to a passive drifter with its aperture pointing downwards, also supported by buoyancy studies and so on. The problem is, of course, the “hook” part and the ascending shaft. I’m not entirely sure. As a personal and absolutely speculative hypothesis without any direct morphological support, I would picture the adults with similar feeding habits as crinoids. The feeble buoyancy provided by the gas in the earlier chambers would make the aperture pointing upwards. They would recline on the muddy floor on their ventral part of the hook, with the tentacles opened and extended mimicking the ones of a crinoid or a spirograph sabellid, which would be used to operate as sedimentation interceptors or suspension feeders. Some forms display spines on the hook (the most extreme example being Anisoceras, for example), which could be used to improve the anchoring. But then again, that might just be wild imagining.
On the response and counter-response for the commented paper. I hadn’t seen them either! To me, Landman and colleague’s response is well exposed and argumented, and focuses on the main concerns of the topic. Especially, I believe that the most important might be “why would both males and females develop hooks?”, which remains unanswered by the subsequent response by Arkhipkin, which, to me, seems a desperate attempt to defend his first hypothesis exploiting the flaws from the former to go back to an initial theory that might have even more defects…
LikeLike
Well, thank you for such a detailed explanation! I was quite puzzled about spikes in ammonoids as well, I hadn’t thought they might play a role in anchoring. It’s nice to see how the hooked hypothesis (silly as it may be) has fuelled this much discussion! (Can’t believe it took me so long to reply, for some reason I thought I had already done it, sorry).
LikeLike
Hello there, just became aware of your blog through Google, and found that it is really informative.
I am gonna watch out for brussels. I will be grateful if you continue this in future.
Many people will be benefited from your writing. Cheers!
LikeLiked by 1 person
Hello there, thanks for reading OnElephantsandBacteria. We are happy that you enjoyed reading our blog. We are continuing slowly, but we are!
LikeLike
I am commenting to let you know what a terrific experience my daughter enjoyed reading through your web page. She noticed a wide variety of pieces, with the inclusion of what it is like to have an awesome helping style to have the rest without hassle grasp some grueling matters.
LikeLike